
After a close aide to Brazil’s president who attended a Mar-a-Lago dinner with Trump tested positive for coronavirus, Trump said he was not "concerned" over the potential exposure.
Man-Made Coronavirus Kills Hundreds (Bill Gates has a Vaccine for That)

It’s been 2 weeks since we first wrote about the novel coronavirus (nCoV-2019) originating in China. Since then, the official numbers have skyrocketed from 630 infections and 17 deaths to over 24,000 infections and roughly 500 deaths.
NaturalNews reports that China is actually keeping two sets of numbers, and that the actual numbers are much higher than the “official” numbers, with over 154,000 infected and almost 25,000 dead. Several nations have evacuated their citizens from china, while airlines have shut down flights to China and other countries.
On January 31st, President Trump declared that the novel coronavirus presents a public health emergency in the United States. That same day, under the guidance of the CDC, the White House Coronavirus Task Force announced a 14-day quarantine for Americans who recently visited Wuhan, China, the epicenter of the coronavirus outbreak.

Over the last two weeks, the Chinese government has quarantined roughly 50 million people in over a dozen cities – the largest quarantine in human history. Cruise liners carrying infected passengers have been denied entry into Japan and passengers will be quarantined on the ships for at least two weeks.
Major manufacturers like Apple and Adidas have shut down their stores and plants in China, and airlines have placed over 30,000 employees on unpaid leave. In the U.S., hundreds of Americans have been evacuated from the Wuhan area and will be immediately quarantined on military bases.
The measures being taken by governments around the world are drastic and unprecedented. The authority that these institutions are willing to assume in the face of a “health emergency” is concerning… and it’s only the beginning.
Social Media Increases Censorship
In the wake of the new virus, Facebook announced that it would remove any content about the coronavirus “with false claims or conspiracy theories that have been flagged by leading global health organizations and local health authorities,” saying such content would violate its ban on misinformation leading to “physical harm.”
This is a much more aggressive policy than Facebook has taken before. Up to this point, censorship efforts have mostly involved restricting search results and advertising while still allowing the content to remain published. The ban will also apply to Instagram, which will subject users to pop-up ads when they click on hashtags related to the virus.
Google has followed suit, pushing updates from the WHO to the top of search results involving the coronavirus. YouTube, owned by Google, will also make it harder to find independent information about the virus, promoting videos from public health organizations and mainstream media outlets.
Twitter said Wednesday that it would begin prompting users who search for the coronavirus to first visit official channels of information about the illness. In the United States, for example, Twitter directs users to the Centers for Disease Control and Prevention, beneath a bold headline that reads: “Know the facts.”
The campaign is running in 15 locations, including the United States, the United Kingdom, Hong Kong, Singapore, and Australia, and “will continue to expand as the need arises,” the company said in a blog post.

Twitter has even banned financial market website Zero Hedge from the social media platform after it published an article linking a Chinese scientist to the outbreak of the fast-spreading coronavirus last week.
Zero Hedge had their account permanently suspended for violating “platform manipulation policy”. The account had 670,000 followers as of its suspension. The suspension was in response to a complaint by Buzzfeed which said Zero Hedge had released the personal information of a scientist from Wuhan in an article that made allegations about coronavirus having been concocted “as a bioweapon.” The article was titled “Is This the Man Behind the Global Coronavirus Pandemic?”
Tik-Tok, a Chinese-owned company, already removes posts and blocks users. This is standard practice in communist China, where state-run media and extreme surveillance of its citizens makes it easy to control what information is released.

The Digital Police State
The effort in China to silence free speech and control the narrative regarding coronavirus has been aggressive. According to a Reuters report:
At least 16 people have been arrested over coronavirus posts in Malaysia, India, Thailand, Indonesia and Hong Kong, while Singapore has used its controversial new “fake news” law, POFMA, to force media outlets and social media users to carry government warnings on their posts and articles saying they contain falsehoods.“Fortunately, we now have POFMA to deal with this fake news,” said Lawrence Wong, one of the ministers heading a Singapore government task force to halt the spread of the virus.At least five people were arrested and released on bail in India’s southwestern state of Kerala over WhatsApp messages, said Aadhithya R, District Police Chief of Thrissur. Six people were arrested in Malaysia on suspicion of spreading false news.In Vietnam, where an army of cyber-censors tracks social media comments for the communist government, at least nine people have been fined and three celebrities asked to explain their actions over posts about coronavirus.Thailand hailed the success of an “anti-fake news centre” it set up last year. Dozens of staff reviewed nearly 7,600 posts in four days from Jan. 25 – leading to 22 posts being highlighted as false on its website and two arrests under computer crimes laws.“The anti-fake news centre is working intensively to verify these rumours and communicating truth to the people,” said Digital Minister Puttipong Punnakanta.
To make matters worse, it turns out that Chinese officials arrested 8 medical professionals who tried to warn people of the disease back in December. Each detainee was part of a medical school’s alumni group on WeChat, a popular social network in China, and they were concerned that SARS, Severe Acute Respiratory Syndrome, was back. (SARS is a type of coronavirus.)
According to The Daily Beast:
It wasn’t long before police detained them. The authorities said these eight doctors and medical technicians were “misinforming” the public, that there was no SARS, that the information was obviously wrong, and that everyone in the city must remain calm. On the first day of 2020, Wuhan police said they had “taken legal measures” against the eight individuals who had “spread rumors.”
Since then, the phenomenal spread of the virus has created cracks even within the normally united front of the Chinese Communist Party. “It might have been fortunate if the public had believed the ‘rumor’ and started to wear masks, carry out sanitization measures, and avoid the wild animal market,” a judge of China’s Supreme People’s Court wrote online last Tuesday.
Li Wenliang, a doctor who was among the eight people who tried to sound the alarm before the coronavirus infected many thousands and killed hundreds, was diagnosed as someone infected with the coronavirus and is being treated at a hospital. Authorities are still actively censoring social-media posts and news articles that question the government’s response to the outbreak. One Wuhan man, Fang Bin, uploaded footage of corpses in a van and a hospital in Wuhan and was then tracked down and taken into custody. His laptop was confiscated, and he had to pedal for three hours on a bicycle to get home after he was questioned, warned, and released. His coronavirus video went viral.
Experts agree that this kind of aggressive censorship could make the virus even more lethal. Leaders within the Chinese government have put their political interests before the good of their people, and the results have been catastrophic thus far.
In recent days, medical experts have found evidence that the origin of the outbreak was not a seafood market in Wuhan, as the Chinese government initially reported. That evidence also suggests that the first human infections occurred in November, if not earlier, rather than in early December.
The true scope of the disease may be worse than we’ve been told. Public officials have been deleting investigative reports by journalists in the area, while many patients who have died of “regular pneumonia” were never tested for coronavirus.
Given the rapid spread of the virus and the enormous economic effects expected, censorship and propaganda are certain to continue — and to extend beyond China’s borders — as the regime seeks to protect its hold on power and international reputation. While Chinese authorities assure domestic and international audiences that their efforts will contain the outbreak, censors are busily deleting social media posts and journalists’ reporting that contradict the official narrative.
An International Conspiracy
Lies and secrecy are common in most communist nations, and China is no exception. It is well-documented that the information released during the SARS outbreak of 2003 was significantly redacted to minimize its true impact.
But what about here in America?
We’ve already reported that, in 2015, a patent was filed by The Pirbright Institute for the live, attenuated coronavirus. The application claims that the new virus could be used to create a vaccine for treating or preventing respiratory viruses. The patent was awarded in 2018.
The Pirbright Institute is funded by the UK Department for Environment, Food, and Rural Affairs, the WHO, and the Bill and Melinda Gates Foundation. All of these entities have been loud supporters of mandatory vaccinations and more government control based on “health concerns.”
And for several years, Bill Gates has been telling us that a pandemic is coming, and in November of 2019, collaborating with the World Economic Forum, the Bill & Melinda Gates Foundation hosted “Event 201” where they ran a simulation of a coronavirus pandemic.
It should come as no surprise that – just yesterday – Bill and Melinda Gates announced that they would be donating $100 million to coronavirus vaccine research and treatment efforts, which were announced as part of the World Health Organization’s (WHO) request for $675 million in global contributions to fight the spread of the disease.
You can bet your bottom dollar that this vaccine will be heavily pushed on all Americans, and probably mandated in some states and cities. But what’s the truth behind the coronavirus?
 The Truth About Coronavirus
The Truth About Coronavirus

Officials would have you believe that the new coronavirus began when someone ate contaminated bat soup at a Wuhan seafood and animal market, but that is a flat-out lie. And a paper published in The Lancet last week has the proof.
The paper, written by a large group of Chinese researchers from several institutions, offers details about the first 41 hospitalized patients who had confirmed infections with what has been dubbed 2019 novel coronavirus (2019-nCoV). In the earliest case, the patient became ill on 1 December 2019 and had no reported link to the seafood market, the authors report.
“No epidemiological link was found between the first patient and later cases,” they state. Their data also show that, in total, 13 of the 41 cases had no link to the marketplace. “That’s a big number, 13, with no link,” says Daniel Lucey, an infectious disease specialist at Georgetown University.
Lucey says if the new data are accurate, the first human infections must have occurred in November 2019—if not earlier—because there is an incubation time between infection and symptoms surfacing. If so, the virus possibly spread silently between people in Wuhan—and perhaps elsewhere—before the cluster of cases from the city’s now-infamous Huanan Seafood Wholesale Market was discovered in late December. “The virus came into that marketplace before it came out of that marketplace,” Lucey asserts.
The much more likely scenario is that a weaponized version of the virus was released (whether intentionally or not) by Wuhan’s Institute of Virology – a level-4 biohazard lab which was studying “the world’s most dangerous pathogens.”
The institute even has an ad for researchers to help use “bats to research the molecular mechanism that allows Ebola and SARS-associated coronaviruses to lie dormant for a long time without causing diseases.”
The job is for a lab run by Dr. Peng Zhou, Ph.D., a researcher at the Wuhan Institute of Virology and Leader of the Bat Virus Infection and Immunization Group. Since 2009, Peng has been the leading Chinese scientist researching the immune mechanism of bats carrying and transmitting lethal viruses in the world.
His primary field of study is researching how and why bats can be infected with some of the most nightmarish viruses in the world including Ebola, SARS and Coronavirus, and not get sick. As part of his studies, Peng also researched mutant Coronavirus strains that overcame the natural immunity of some bats; these are “superbug” Coronavirus strains, which are not resistant to any natural immune pathway, and now appear to be out in the wild.
The institute is about 13 miles away from the market that China claims as the source of the outbreak.
A study by 5 Greek scientists (published 1/27/2020) examined the genetic relationships of nCoV-2019 and found that “the new coronavirus provides a new lineage for almost half of its genome, with no close genetic relationships to other viruses within the subgenus of sarbecovirus,” and has an unusual middle segment never seen before in any coronavirus.
What exactly does that mean? Basically, this means that we are dealing with a brand new type of ‘man-made’ coronavirus. The study’s authors rejected the original hypothesis that the virus originated from random natural mutations between different coronaviruses.
Dany Shoham, a former Israeli military intelligence officer who has studied Chinese biological warfare, has also linked the virus to China’s covert biological weapons program. Mr. Shoham holds a doctorate in medical microbiology. From 1970 to 1991, he was a senior analyst with Israeli military intelligence for biological and chemical warfare in the Middle East and worldwide.
“Certain laboratories in the institute have probably been engaged, in terms of research and development, in Chinese [biological weapons], at least collaterally, yet not as a principal facility of the Chinese BW alignment,” Mr. Shoham told The Washington Times. Work on biological weapons is conducted as part of dual civilian-military research and is “definitely covert,” he said.
In a sad turn of events, the Chinese doctor who tried to warn others about the Wuhan coronavirus, has died. Li Wenliang, a 34-year-old doctor working in Wuhan, raised the alarm about the novel coronavirus on December 30th. Soon after he posted the message, Li was accused of rumor-mongering by the Wuhan police. He was one of several medics targeted by police for trying to blow the whistle on the deadly virus in the early weeks of the outbreak. Li was hospitalized on January 12th after contracted the virus from one of his patients, and he was confirmed to have nCoV-2019 on February 1st.


The question is … If the Chinese made this virus in a lab, what was their purpose? And why did they arrest the doctor who tried to inform the rest of the world?
The world needs an answer.
Wolves in Sheep’s Clothing
As long as we continue to believe the lies promulgated by communist state-run media, we’re going to stay in the dark. Government agencies around the world have been happy to sensationalize the virus as an excuse for extreme control over their citizens.
People are being detained and quarantined. Travel and business have been seriously disrupted. The global economy is in turmoil. Scientists and journalists are being silenced and even arrested. And social media companies founded in the freest country on earth are deleting any content that suggests that this is a man-made problem.
People are scared. Tens of millions have had their lives put on hold. Hundreds have died in the last week.
But don’t worry… be healthy! There are herbs and natural substances that can have a profound effect on viruses and your immune function.
Surprise surprise! Bill Gates, the WHO, and everyone else in the medical industry are working on a nCoV-2019 vaccine … a vaccine that they patented half a decade before the virus appeared … a virus that was created in a lab.
But that’s just a “coincidence,” right?
Here’s another incredible “coincidence” …
For several years, Bill Gates has been telling us that a pandemic is coming, and on October 18, 2019 (collaborating with the World Economic Forum), the Bill & Melinda Gates Foundation hosted “Event 201” where they ran a simulation of a coronavirus pandemic.
A meeting of the “globalist health minds” occurred on November 6th to discuss its findings with 9 recommendations.
Surprise surprise! One recommendation, of course, included deleting/erasing/censoring “disinformation” in the WHO’s eyes.
Coronavirus & HIV – The Truth?
The conspiracy network is alive and thriving and the gist of this is that the coronavirus is created in some lab because it has a sequence that is similar to HIV. I have two very good sources and they point out that this claim is MOST LIKELY completely bogus. Why? Because this very same sequence also exists in many other viruses and it is NOT unique to the coronavirus or HIV, which by the way was a virus.
This claim comes from a questionable scientist based in India where he took the genome sequences of Wuhan coronavirus, without any peer review, and claimed that it had similarities to the genome of…HIV. Some have argued this is why it is even spreading so fast. Sorry, that is because of how it is transmitted which is like the seasonal flu whereas HIV was transmitted predominantly from sexual contact or a blood transfusion with tainted blood.
Reliable sources found 4 insertions in the spike glycoprotein (S) which are unique to the 2019-nCoV and are not present in other coronaviruses. Most interestingly, amino acid residues in all the 4 inserts have identity or similarity to those in the HIV-1 gp120 or HIV-1 Gag.
Before we jump to the next zombie apocalypse, I question why any reliable scientist would spurt out such findings without peer-review? Everything is turning on this distrust of China. If the coronavirus was spliced in a lab by them for military purposes, then surely they would not hand the evidence over to the rest of the world. Let’s be rational here.
Given that the Wuhan coronavirus (2019-nCoV) appears to have four additions to its genome that similar viruses, like SARS and MERS, are lacking, is not proof in and of itself. This implies a lower death-rate, but perhaps a higher contagion rate being more similar to the seasonal flu. These additions being touted by the scientist in India claim they are identical or extremely similar to parts of the HIV genome. Many go on to speculate that this may be why the coronavirus is spreading so quickly, and also that something sinister may be afoot. He did not bother to elaborate on what methods were used to genetically insert these into the coronavirus.
Sources checking whether it was true that HIV and coronavirus have similarities found that while these sequences do pop up in HIV, they ALSO pop up in so many other viruses. There’s absolutely NO EVIDENCE that they are immediately suspect or that they were somehow genetically spliced in by the lab in China which is the implication. One out of four sequences is indeed found in both Wuhan coronavirus and HIV. The same sequence also appears in a virus that infects Streptococcus (spherical bacteria), which is of a rat virus in origin. They also appear in what is known as an “acute bee paralysis virus.”
Sorry, perhaps the culprit may just be mother nature and not some mad scientist in a lab. We should wait for a peer-to-peer review, but this claim from India appears to be bogus on the surface. It is not pointing out that this sequence also appears in many other viruses. That is really misleading a lot of people in the conspiracy world who seem to want to bring about a horror move like some zombie apocalypse.
New coronavirus emerges from bats in China, devastates young swine
Identified in same region, from same bats, as SARS coronavirus
- April 4, 2018
- NIH/National Institute of Allergy and Infectious Diseases
- A newly identified coronavirus that killed nearly 25,000 piglets in 2016-17 in China emerged from horseshoe bats near the origin of the severe acute respiratory syndrome coronavirus (SARS-CoV), which emerged in 2002 in the same bat species. The new virus, called swine acute diarrhea syndrome coronavirus (SADS-CoV), doesn't appear to infect people, unlike SARS-CoV.
FULL STORY
A newly identified coronavirus that killed nearly 25,000 piglets in 2016-17 in China emerged from horseshoe bats near the origin of the severe acute respiratory syndrome coronavirus (SARS-CoV), which emerged in 2002 in the same bat species. The new virus is named swine acute diarrhea syndrome coronavirus (SADS-CoV). It does not appear to infect people, unlike SARS-CoV which infected more than 8,000 people and killed 774. No SARS-CoV cases have been identified since 2004. The study investigators identified SADS-CoV on four pig farms in China's Guangdong Province. The work was a collaboration among scientists from EcoHealth Alliance, Duke-NUS Medical School, Wuhan Institute of Virology and other organizations, and was funded by the National Institute of Allergy and Infectious Diseases, part of the National Institutes of Health. The research is published in the journal Nature.
The researchers say the finding is an important reminder that identifying new viruses in animals and quickly determining their potential to infect people is a key way to reduce global health threats.
SADS-CoV began killing piglets on a farm near Foshan in Guangdong Province in late October 2016. Investigators initially suspected porcine epidemic diarrhea virus (PEDV) as the cause. PEDV is a type of coronavirus common to swine that had been identified at the Foshan farm. Detection of PEDV ceased by mid-January 2017, yet piglets continued to die, suggesting a different cause. Scientists say separating sick sows and piglets from the rest of the herd helped stop the outbreak of SADS-CoV by May 2017.
Investigators confirmed the connection of SADS-CoV to bats by identifying the new virus in the small intestine of piglets from the outbreak. They then determined that the genetic sequence of SADS-CoV is similar to that of a bat coronavirus discovered in 2007 and looked for evidence of SADS-CoV in bat specimens collected from 2013 to 2016 in Guangdong Province. The new virus appeared in 71 of 596 specimens (11.9 percent).
The researchers also tested 35 farm workers who had close contact with sick pigs, none of whom tested positive for SADS-CoV.
Currently six coronaviruses are known to cause disease in people, but so far only two of them -- SARS-CoV and Middle East Respiratory Syndrome coronavirus -- have caused large outbreaks of fatal illness in people.
This research was supported by NIAID award R01AI110964.
Review
Bat Coronaviruses in China


1
CAS Key Laboratory of Special Pathogens and Biosafety, Wuhan Institute of Virology, Chinese Academy of Sciences, Wuhan 430071, China
2
University of Chinese Academy of Sciences, Beijing 100049, China
*
Author to whom correspondence should be addressed.
Received: 29 January 2019 / Accepted: 26 February 2019 / Published: 2 March 2019
Abstract
:
During the past two decades, three zoonotic coronaviruses have been identified as the cause of large-scale disease outbreaks–Severe Acute Respiratory Syndrome (SARS), Middle East Respiratory Syndrome (MERS), and Swine Acute Diarrhea Syndrome (SADS). SARS and MERS emerged in 2003 and 2012, respectively, and caused a worldwide pandemic that claimed thousands of human lives, while SADS struck the swine industry in 2017. They have common characteristics, such as they are all highly pathogenic to humans or livestock, their agents originated from bats, and two of them originated in China. Thus, it is highly likely that future SARS- or MERS-like coronavirus outbreaks will originate from bats, and there is an increased probability that this will occur in China. Therefore, the investigation of bat coronaviruses becomes an urgent issue for the detection of early warning signs, which in turn minimizes the impact of such future outbreaks in China. The purpose of the review is to summarize the current knowledge on viral diversity, reservoir hosts, and the geographical distributions of bat coronaviruses in China, and eventually we aim to predict virus hotspots and their cross-species transmission potential.
Keywords:
coronavirus; bat; epidemiology; cross-species; zoonosis1. Introduction
Fifteen years after the first highly pathogenic human coronavirus caused the severe acute respiratory syndrome coronavirus (SARS-CoV) outbreak, another severe acute diarrhea syndrome coronavirus (SADS-CoV) devastated livestock production by causing fatal diseases in pigs. Both outbreaks began in China and were caused by coronaviruses of bat origin [1,2]. This increased the urgency to study bat coronaviruses in China to understand their potential of causing another virus outbreak.
In this review, we collected information from past epidemiology studies on bat coronaviruses in China, including the virus species identified, their host species, and their geographical distributions. We also discuss the future prospects of bat coronaviruses cross-species transmission and spread in China.
2. Why Study Bat Coronaviruses in China?
2.1. Coronavirus Taxonomy
Coronaviruses (CoVs) belong to the subfamily Orthocoronavirinae in the family Coronaviridae and the order Nidovirales. CoVs have an enveloped, crown-like viral particle from which they were named after. The CoV genome is a positive-sense, single-strand RNA (+ssRNA), 27–32 kb in size, which is the second largest of all RNA virus genomes. Typically, two thirds of the genomic RNA encodes for two large overlapping polyproteins, ORF1a and ORF1b, that are processed into the viral polymerase (RdRp) and other nonstructural proteins involved in RNA synthesis or host response modulation. The other third of the genome encodes for four structural proteins (spike (S), envelope (E), membrane (M), and nucleocapsid (N)) and other accessory proteins. While the ORF1a/ORF1b and the four structural proteins are relatively consistent, the length of the CoV genome is largely dependent on the number and size of accessory proteins [3].
Compared with other RNA viruses, the expanded genome size of CoVs is believed to be associated with increased replication fidelity, after acquiring genes encoding RNA-processing enzymes [4]. Genome expansion further facilitates the acquisition of genes encoding accessory proteins that are beneficial for CoVs to adapt to a specific host [5]. As a result, genome changes caused by recombination, gene interchange, and gene insertion or deletion are common among CoVs. The CoV subfamily is expanding rapidly, due to the application of next generation sequencing which has increased the detection and identification of new CoV species. As a result, CoV taxonomy is constantly changing. According to the latest International Committee of Taxonomy of Viruses (ICTV) classification, there are four genera (α-, β-, δ-, and γ-) consisting of thirty-eight unique species in the subfamily [6]. The number of species will continue to increase, as there are still many unclassified CoVs [7,8].
CoVs cause disease in a variety of domestic and wild animals as well as in humans, where α- and β-CoVs mainly infect mammals and γ- and δ-CoVs mainly infect birds (Table 1). Two highly pathogenic β-CoVs, SARS-CoV, and MERS-CoV have caused pandemics in humans since 2002 [1,9]. Originating in China and then spreading to other parts of the world, SARS-CoV infected around 8000 individuals with an overall mortality of 10% during the 2002–2003 pandemic [1]. Since its emergence in 2012 in the Middle East, MERS-CoV spread to 27 countries, resulting in 2249 laboratory-confirmed cases of infection with an average mortality of 35.5% (until September 2018) [9]. Besides these two viruses, α-CoVs 229E and NL63 and β-CoVs OC43 and HKU1 can also cause respiratory diseases in humans [10]. Moreover, CoVs cause pandemic disease in domestic and wild animals (Table 1). SADS-CoV was recently identified as the etiological agent responsible for a large-scale outbreak of fatal disease in pigs in China that caused the death of more than 20,000 piglets [2]. Porcine epidemic diarrhea virus (PEDV) and transmissible gastroenteritis virus (TGEV) that belong to α-CoV and porcine δ-CoV (PDCoV) are also important emerging and re-emerging viruses in pigs that pose significant economic threat to the swine industry [11]. In addition, avian infectious bronchitis virus (IBV, γ-CoV) causes a highly contagious disease that affects poultry production worldwide [12]. Coronaviruses have also been associated with catarrhal gastroenteritis in mink (MCoV) and whale deaths (BWCoV-SW1) [13,14].

2.2. Linking Bats to Coronaviruses
Bat are the only mammals with the capability of powered flight, which enables them to have a longer range of migration compared to land mammals. Bats are also the second largest order of mammals, accounting for about a fifth of all mammalian species, and are distributed worldwide. Phylogenetic analysis classified bats into two large suborders—the Yinpterochiroptera, consisting of one Pteropodidae (megabat) and five Rhinolophoidea (microbat) families, and the Yangochiroptera comprising a total of thirteen microbat families [15].
It is hypothesized that flight provided the selection pressure for coexistence with viruses, while the migratory ability of bats has particular relevance in the context of disease transmission [16]. Indeed, bats were linked to a few highly pathogenic human diseases, supporting this hypothesis. Some of these well characterized bat viruses, including bat lyssaviruses (Rabies virus), henipaviruses (Nipah virus and Hendra virus), CoVs (SARS-CoV, MERS-CoV, and SADS-CoV), and filoviruses (Marburg virus, Ebola virus, and Mengla virus), pose a great threat to human health [16,17]. A comprehensive analysis of mammalian host–virus relationships demonstrated that bats harbor a significantly higher proportion of zoonotic viruses than other mammalian orders [18]. Viruses from most of the viral families can be found in bats [16].
Bats are now recognized as important reservoir hosts of CoVs (Table 1). Although civet cats were initially identified as the animal origin of SARS-CoV, bats were soon found to be the most likely natural reservoir hosts of this virus [19,20,21]. Long-term surveillance revealed an average 10% SARS-related CoV nucleotide positivity in bats, including some viruses that can use same human entry receptor ACE2 as SARS-CoV [7,22]. Similarly, bats have been proposed to harbor the progenitor viruses of MERS-CoV, although dromedary camels can transmit this virus to humans directly [9]. The most recent SADS-CoV spillover was traced back to bats [2]. In addition, bats also carry α-CoVs that are related to pathogenic human 229E- and NL63-CoVs, as well as pandemic swine coronavirus PEDV [23,24]. In summary, bats carry major α- (10 out of 17) and β- (7 out of 12) CoV species that may spillover to humans and cause disease (Table 1). Attributed to the wide distribution of bats, CoVs can be found worldwide, including China [25].
2.3. Why China?
Two bat CoVs caused outbreaks in China; it is thus urgent to study the reasons to avoid future outbreaks. China is the third largest territory and is also the most populous nation in the world. A vast homeland plus diverse climates bring about great biodiversity including that of bats and bat-borne viruses—most of the ICTV coronavirus species (22/38) were named by Chinese scientists studying local bats or other mammals. The majority of the CoVs can be found in China (Table 1). Moreover, most of the bat hosts of these CoVs live near humans, potentially transmitting viruses to humans and livestock. Chinese food culture maintains that live slaughtered animals are more nutritious, and this belief may enhance viral transmission.
It is generally believed that bat-borne CoVs will re-emerge to cause the next disease outbreak. In this regard, China is a likely hotspot. The challenge is to predict when and where, so that we can try our best to prevent such outbreaks.
3. Bat Coronaviruses That Are Associated with Diseases
3.1. SARS-Related Coronaviruses
In November 2012, the first case of SARS was recorded in Foshan city, Guangdong Province, China (Figure 1). In 2005, two independent Chinese groups reported the first bat SARS-related CoV (SARSr-CoV) that was closely related to human SARS-CoV, implying a bat origin of the latter [20,21]. Since then, more bat SARSr-CoV isolates were identified in China (Table 1). Genome identities of these bat SARSr-CoVs are as high as 92% to human SARS-CoV, but their major receptor binding spike proteins cannot use the human virus entry receptor ACE2 [67]. Whether they are the progenitor viruses of SARS-CoV is debatable. In 2013, the isolation of a bat SARSr-CoV that uses the ACE2 receptor provided the strongest evidence of the bat origin of SARS-CoV [22]. Furthermore, the building blocks for SARS-CoV were identified from eleven different SARSr-CoV viral strains in a five-year surveillance program in a cave inhabited by multiple species of horseshoe bats in Yunnan Province, China [62].

Figure 1. Geographical distribution of bat coronaviruses (CoVs) and their corresponding bat hosts in China. Each red box represents one CoV positive sample found in that particular bat species. One dot matrix was drawn for each province where a CoV positive sample had been reported. Guangdong Province, where SARS and SADS began, is circled in red. Abbreviations of bat species and virus species are indicated.
SARSr-CoVs found in China show great genomic diversity (Figure 2). Sequence identities of the conserved 440 bp RdRp region ranges from 80 to 100% with human SARS-CoV. CoV diversity in bats is thought to be shaped by both species richness and geographical distribution, and CoVs exhibit clustering at the bat genera level, with these genus-specific clusters largely associated with distinct CoV species [25]. Our analysis supports this theory. SARSr-CoVs are present in different bat species but all belong to the family of Rhinolophidae and Hipposideridae (Figure 1). Chaerephon plicata bats were also reported as carriers in one study, but this cannot be conclusively supported without molecular identification of the bat species [8]. In China, horseshoe bat species (Rhinolophus spp.) are widely distributed, including R. sinicus, R. ferrumequinum, R. macrotis, R. pearsoni, and R. pusillus, and are also the most frequent SARSr-CoV carriers throughout the nation [7,8,20,21,22,27,40,43,45,58,59,61,62,63,68] (Figure 1). The most variable regions among bat SARSr-CoVs are the S and ORF8 genes [62]. The S protein in certain strains is capable of using human ACE2 as a receptor and thus poses a direct threat to humans [69]. Interestingly, all the SARSr-CoVs that are capable of using human ACE2 were found in R. sinicus in Yunnan Province [7,22,27,62]. Other SARSr-CoVs that cannot use human ACE2 were distributed in multiple provinces, from north Jilin, Shaanxi, Shanxi to south Hubei, Zhejiang, Yunnan, Guizhou, and Guangdong (Figure 1). Another protein, ORF8, was suggested to be important for interspecies transmission, as most human SARS-CoV epidemic strains contain a signature 29-nucleotide deletion in ORF8 compared to civet SARSr-CoVs, which results in the formation of two separate open reading frames, ORF 8a and 8b [40]. Only two R. ferrumequinum and one R. sinicus from Yunnan Province carried viruses that possess ORF8 proteins with exceptionally high amino acid identities to that of human/civet SARSr-CoVs [40,62]. It was strongly suggested that SARS-CoV most likely originated from Yunnan Rhinolophus bats via recombination events among existing SARSr-CoVs.

Figure 2. Genetic diversity of bat CoV in China. Sequences of 440 bp conserved the viral polymerase (RdRp) region for each CoV species were compared to related reference sequences. Reference genomes used: BtCoV-HKU10, NC_018871.1; BtRfCoV-HuB13, NC_028814.1; BtMiCoV-1, EU420138.1; BtMiCoV-HKU8, NC_010438.1; BtRhCoV-HKU2, MF094682.1; BtHpCoV-ZJ13, NC_025217.1; MERSr-CoV, NC_038294.1; BtPiCoV-HKU5, NC_009020.1; BtTyCoV-HKU4, NC_009019.1; BtRoCoV-GCCDC1, MG762606.1; BtRoCoV-HKU9, NC_009021.1; and SARSr-CoV, NC_004718.3. Notably, samples that were positive for BtMy-Sax11, BtNy-Sc13, and BtScCoV-512 were also identified in China. These were not taken into account here as too few sequences were available.
These studies revealed that various SARSr-CoVs capable of using human ACE2 are still circulating among bats in China, highlighting the possibly of another SARS-like disease outbreak. Certain areas in Yunnan Province are hotspots for spillover. To support this hypothesis, we provide serological evidence of bat SARSr-CoV infection in humans in Yunnan Province where no prior exposure to SARS-CoV was recorded [70]. The majority of the SARSr-CoVs appear not able to use ACE2, but their infectivity or pathogenesis to humans are still unknown. Frequent interspecies recombination may result in another human infectious coronavirus from these SARSr-CoVs. Furthermore, there are still unanswered questions about SARS, e.g., ‘Why did the first SARS case occur in Guangdong Province, but all the human-ACE2-using SARSr-CoVs were found in Yunnan Province?’ and ’Why does R. sinicus in certain areas carry human-ACE2-using SARSr-CoVs but no other Rhinolophus species carry the same viruses?’ Above all, further extensive surveillance of SARSr-CoVs in China is warranted.
3.2. MERS-Cluster Coronaviruses
Different to bat SARSr-CoV, MERS-cluster CoVs were found in bats before the MERS disease outbreaks. Two bat CoVs, Tylonycteris HKU4 and Pipistrellus HKU5 were first described as putative group 2c CoVs in 2006 in China. They were associated with the HCoV-EMC (MERS-CoV) that started the 2012 pandemic [9,38,39]. It is generally accepted that Middle East dromedary camels were the major animal source for the zoonotic transmission of human MERS, while bats harbor CoVs that shared common ancestry with MERS-CoV [71]. Extensive global surveys revealed a wide distribution of largely diverged MERS-cluster CoVs (lineage 2c CoVs) [71]. Two closely related Neoromicia zuluensis bat CoVs, NeoCoV and PREDICT/PDF-2180, were subsequently found, further supporting the idea that MERS-CoV was descended from an ancestral virus of African bats [72,73]. So far, three species of lineage 2c CoVs have been found in bats, according to the latest CoV taxonomy reports. Based on phylogenetic trees constructed using RdRp, ORF1, S1, and N sequences, bat MERS-related CoVs (MERSr-CoVs) are the closest relatives of MERS-CoV, followed by HKU4-CoV and HKU5-CoV. However, in the S1 region, MERS-CoV was much closer to HKU4-CoV than to MERSr-CoV or HKU5-CoV. Likewise, pseudovirus assays showed that the MERSr-CoV (HKU25 and 422CoV) spike protein can use human DPP4 for entry into hDPP4-expressing cells, although with lower efficiency than that of MERS-CoV or HKU4-CoV spike proteins [49,50]. There is no evidence of HKU5-CoV using the human DPP4 receptor [74].
All three types of bat MERS-cluster CoVs can be found in China (Figure 1 and Figure 2). Their reservoir hosts all belong to the Vespertilionidae family. MERSr-CoV can be found in multiple bat species, including Pipistrellus bats (P. abramus and P. pipistrellus), great evening bats (Ia io), particolored bats (Vespertilio superans), and Chinese pipistrelle bats (Hypsugo pulveratus) [49,50,52]. Due to this wide host spectrum, MERSr-CoV also showed a large genetic diversity, ranging from 72 to 100% in the conserved 440 bp RdRp region. In contrast, HKU4-CoVs were only carried by Tylonycteris bats (T. pachypus and T. robustula) and were relatively conserved [38,39,49] (Figure 2). HKU5-CoVs were found in different Pipistrellus bats (P. abramus, P. pipistrellus, P. minus, and P. spp.) [8,36,38,39,49,51]. Like HKU4-CoVs, they are also relatively conserved. The range of distribution varies, depending on MERS-cluster CoV species. HKU5-CoVs should be the most widely distributed CoVs among the three as their hosts, Pipistrellus bats, live close to humans. However, the reported CoV positive samples can only be found in Guangdong, Hong Kong, and Macau, possibly due to a lack of investigation in other provinces. In contrast, MERSr-CoVs were reported in multiple bat species in Sichuan, Guangdong, and Hong Kong at a much lower level than HKU5-CoVs. Similarly, Tylonycteris bats are a rare bat species that live in bamboo, which restricted the distribution of HKU4-CoVs to certain locations in Guangdong, Guangxi, Yunnan, Guizhou, Hong Kong, and Macau (Figure 1). To sum up, it appears that the risk of MERS-cluster CoV spillover to humans leading to an epidemic in China is low for the following reasons: (1) the geographical distribution of MERSr-CoVs and HKU4-CoVs that have the potential to infect humans (capable of using human entry receptors) is limited, and (2) HKU5-CoVs that widely exist in Chinese bats across the nation have not obtained the ability of using human entry receptors. However, we should not underestimate the possibility of recombination among different bat CoVs that lead to the generation of potential pandemic viruses.
3.3. HKU2 (SADS)-Related CoV (HKU2r-CoV)
HKU2r-CoVs have only been reported in China and Kenya. From studies in China, HKU2r-CoVs have been frequently found in Rhinolophus bats (R. affinis, R. sinicus, R. rex, and R. pusillus) in several provinces before the SADS outbreak [2,7,8,38,41,44]. So far, the virus has been reported in Hong Kong, Guangdong, Yunnan, and Tibet. There are perhaps more to be discovered in other provinces considering the wide range of Rhinolophus bats. Notably, these bat species, which constantly interact with both livestock and humans in China, also harbor SARSr-CoVs (see Section 3.1). Likewise, HKU2r-CoVs showed a high genetic diversity with SARSr-CoVs (Figure 2). Due to these characteristics, HKU2r-CoVs were listed as viruses that were highly likely to cross species to humans. The novel HKU2r-CoV, swine acute diarrhea syndrome coronavirus (SADS-CoV), was identified as the etiological agent responsible for a large-scale outbreak of fatal disease in pigs in China, Guangdong Province in 2017 [2]. The entry receptor of SADS-CoV has not been identified, yet this virus showed a capacity for infecting a wide range of human, swine, and bat cells (unpublished data). In China, the high density of pig farms and the wide distribution of host bat species promote the possibility of future HKU2r-CoV cross-species transmission [75]. Thus, studies on bat HKU2r-CoVs spillover potential and their pathogenesis are urgent.
4. A SADS-CoV Model of Prediction and Other Hotspot Viruses
To predict the next CoV that will cause a virus outbreak in future, we list the general factors that may contribute to this outbreak. Firstly, bats host a large number of highly diverse CoVs. It is known that CoV genomes regularly undergo recombination during infection, and a rich gene pool can facilitate this process. Secondly, bat species are widely distributed and live close to humans. Thirdly, the viruses are pathogenic and transmissible. In this context, SADS-CoV and SARS-CoV outbreaks in China are not unexpected. By this model, there are other CoVs that have not yet caused virus outbreaks but should be monitored.
Within the family Vespertilionidae, the mouse-eared bats (Myotis) which favor roosting in abandoned human facilities are also a widespread genus of bats besides Pipistrellus bats. They carry a large number and genetically diversified HKU6-CoVs that are closely related to Myotis ricketti α-CoV Sax-2011 [36,38]. Moreover, bent-winged bats (Miniopterus spp.) carry a large variety of α-CoVs. One of the most frequently detected viruses is HKU8-CoV, which was first described circulating in M. pusillus in Hong Kong in 2005. Later, it was also found in M. magnate, M. fuliginosus, and M. schreibersii in Hong Kong, Guangdong, Yunnan, Fujian, and Hubei provinces, showing a great genetic diversity [32,33,34,35,37,41,60] (Figure 1). Besides HKU8-CoVs, bent-winged bats (Miniopterus spp.) also harbor a large amount of Miniopterus bat CoV 1 (BtMiCoV-1), which were called CoV1A or CoV1B previously. This viral species was found almost as frequently as HKU8-CoV in multiple provinces in China in Miniopterus bats, although these viruses showed a relatively small sequence variation between each other [32,33,34,35,37,41,60]. Genetic analysis indicates that BtMiCoV-1, HKU8-CoV, and HKU7-CoV (previous name) are different but closely related CoVs circulating in bent-winged bats and may have descended from a common ancestor [34]. Additionally, Rousettus leschenaultii bats in the family of Pteropodidae harbor HKU9-CoVs. As a fruit bat, Rousettus leschenaultii has a wider flying range than most of the insectivorous bats in China, thus it may carry viruses over long distances. A comparison of the reported HKU9-CoV sequences showed a high genetic diversity within this viral species [55,56,57] (Figure 2). The last CoV that should be mentioned is HKU10-CoV. HKU10-CoVs can be found in bats from different genera (Rousettus leschenaultii and Hipposideros pomona), suggesting interspecies transmission between bats [7,26,27,39]. A genetic difference can also be observed for this virus species (Figure 2). Above all, these viruses fit well in our SADS prediction model and should be monitored in our future studies.
5. Other Bat CoVs in China
In 2016, a novel β-CoV, Ro-BatCoVGCCDC1, was identified from the Rousettus leschenaultii bat. However, we confirmed the host was a closely related Eonycteris spelaea bat upon species identification and then renamed the virus as BtEoCoV-GCCDC1 (Table 1). The uniqueness of this virus is that it contains a gene that most likely originated from the p10 gene of a bat orthoreovirus [53]. A two-year follow-up study also illustrated that BtEoCoV-GCCDC1 persistently circulates among bats. Different to the genetically diverged HKU9-CoV, this virus is highly conserved (Figure 2). BtEoCoV-GCCDC1 has only been found in south Yunnan Province so far [54,55]. In addition, there are other bat CoVs that have been identified in China: Rhinolophus ferrumequinum α-CoV HuB-2013 [8], Myotis ricketti α-CoV Sax-2011 [8,37], Nyctalus velutinus α-CoV SC-2013 [8], Scotophilus bat CoV 512 [37], Hipposideros bat β-CoV Zhejiang2013, and a Murina leucogaster bat CoV, which has been described as the evolutionary ancestor of PEDV [37]. Notably, there are still many unclassified bat CoVs circulating in China, particularly in the northern part of the nation where bat viruses were rarely studied (Figure 1). According to the criteria defined by the ICTV, the CoV family will most likely expand following further investigation of bat CoVs in China.
6. Coexistence of Different Coronaviruses or Other Viruses in Bats
The coexistence of more than two viruses in the same bat is quite common for some bat species. The coexistence of Miniopterus bat CoV 1 and HKU8-CoV in one bat has been frequently reported [7,34]. Another example is the coexistence between Rhinolophus HKU2-CoVs (SADS-CoV) and SARSr-CoVs that caused the virus outbreaks, respectively [2,45]. Real-time monitoring this bat genus is necessary for the prevention of future SARS-like outbreaks. Moreover, two or more distinct genotypes of HKU9-CoVs were reported to coexist in a single Rousettus bat [56]. The coexistence of HKU9-CoVs and a new identified bat filovirus (Mengla virus) that is phylogenetically related to the Ebola and Marburg viruses was also identified from Rousettus bats [17,55]. Given that a bat orthoreovirus p10 gene was incorporated in the BtEoCoV-GCCDC1 genome, recombination between the bat filovirus and HKU9-CoV cannot be excluded. Other pairs were also recorded—HKU8-CoV with unclassified α-CoV [7], HKU2-CoV with unclassified α-CoV [7], HKU10-CoV with unclassified β-CoV [7], and HKU6-CoV with bat adenovirus [36].
7. Conclusions
Two bat origin CoVs caused large-scale epidemics in China over fourteen years, highlighting the risk of a future bat CoV outbreak in this nation. In this review, we have summarized the current findings related to bat CoV epidemiology in China, aiming to explore the associations between CoV species, bat species, and geographical locations, and eventually we aim to predict the cross-species transmission potential of these bat CoVs. Admittedly, the analysis may be affected by inaccurate or incomplete data. For example, not all research groups performed bat species identification or used Global Positioning System (GPS) during bat sampling. Bats in the north or west provinces were not surveyed either. Nonetheless, we believe this analysis is a good starting point for further research. Moreover, there are other outstanding questions that should be addressed in future studies: (1) given that most of the ICTV classified CoV species are from bats, why there are so many genetically divergent CoVs in bats, (2) the pathogenesis of most bat CoVs in humans remains unknown as the viruses have never been isolated or rescued—apart from the viruses identified during the outbreaks, many viruses pose a threat to human health, (3) although SARS-CoV and SADS-CoV were known to be transmitted from bats to human or swine, their exact transmission routes are unknown, and (4) why bats can maintain CoVs long-term without showing clinical symptoms of diseases. A unique bat immunity model has been proposed. The authors have shown that constitutively expressed bat interferon α may protect bats from infection [76], while some particularly dampened immune pathways may allow bats to have a higher tolerance against viral diseases [77]. While we start to unveil the mystery of unique bat immunity, there is still long way to go before we can fully understand the relationship between bats and coronaviruses.
Author Contributions
P.Z. and Z.-L.S. designed the study. Y.F. and K.Z. analyzed the data. P.Z. and Y.F. prepared the manuscript.
Funding
The work was supported by China’s National Science and Technology Major Project on Infectious Diseases (2018ZX10101004) and the National Natural Science Foundation of China (Excellent Young Scholars to PZ 81822028 and 81661148058).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Drosten, C.; Gunther, S.; Preiser, W.; van der Werf, S.; Brodt, H.R.; Becker, S.; Rabenau, H.; Panning, M.; Kolesnikova, L.; Fouchier, R.A.M.; et al. Identification of a novel coronavirus in patients with severe acute respiratory syndrome. N. Engl. J. Med. 2003, 348, 1967–1976. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Fan, H.; Lan, T.; Yang, X.L.; Shi, W.F.; Zhang, W.; Zhu, Y.; Zhang, Y.W.; Xie, Q.M.; Mani, S.; et al. Fatal swine acute diarrhoea syndrome caused by an HKU2-related coronavirus of bat origin. Nature 2018, 556, 255. [Google Scholar] [CrossRef] [PubMed]
- Coronavirinae in ViralZone. Available online: https://viralzone.expasy.org/785 (accessed on 28 January 2019).
- Subissi, L.; Posthuma, C.C.; Collet, A.; Zevenhoven-Dobbe, J.C.; Gorbalenya, A.E.; Decroly, E.; Snijder, E.J.; Canard, B.; Imbert, I. One severe acute respiratory syndrome coronavirus protein complex integrates processive RNA polymerase and exonuclease activities. Proc. Natl. Acad. Sci. USA 2014, 111, E3900–E3909. [Google Scholar] [CrossRef] [PubMed]
- Forni, D.; Cagliani, R.; Clerici, M.; Sironi, M. Molecular Evolution of Human Coronavirus Genomes. Trends Microbiol. 2017, 25, 35–48. [Google Scholar] [CrossRef] [PubMed]
- ICTV Virus Taxonomy: 2018 Release. 2018. Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 28 January 2019).
- Ge, X.Y.; Wang, N.; Zhang, W.; Hu, B.; Li, B.; Zhang, Y.Z.; Zhou, J.H.; Luo, C.M.; Yang, X.L.; Wu, L.J.; et al. Coexistence of multiple coronaviruses in several bat colonies in an abandoned mineshaft. Virol. Sin. 2016, 31, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Yang, L.; Ren, X.; He, G.; Zhang, J.; Yang, J.; Qian, Z.; Dong, J.; Sun, L.; Zhu, Y.; et al. Deciphering the bat virome catalog to better understand the ecological diversity of bat viruses and the bat origin of emerging infectious diseases. ISME J. 2016, 10, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Zaki, A.M.; van Boheemen, S.; Bestebroer, T.M.; Osterhaus, A.D.M.E.; Fouchier, R.A.M. Isolation of a Novel Coronavirus from a Man with Pneumonia in Saudi Arabia. N. Engl. J. Med. 2012, 367, 1814–1820. [Google Scholar] [CrossRef] [PubMed]
- Graham, R.L.; Donaldson, E.F.; Baric, R.S. A decade after SARS: Strategies for controlling emerging coronaviruses. Nat. Rev. Microbiol. 2013, 11, 836–848. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Yoo, D. Immune evasion of porcine enteric coronaviruses and viral modulation of antiviral innate signaling. Virus Res. 2016, 226, 128–141. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, H.; Zhao, J.; Zhong, Q.; Jin, J.H.; Zhang, G.Z. Evolution of infectious bronchitis virus in China over the past two decades. J. Gen. Virol. 2016, 97, 1566–1574. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Mihindukulasuriya, K.A.; Wu, G.; St Leger, J.; Nordhausen, R.W.; Wang, D. Identification of a novel coronavirus from a beluga whale by using a panviral microarray. J. Virol. 2008, 82, 5084–5088. [Google Scholar] [CrossRef] [PubMed]
- Vlasova, A.N.; Halpin, R.; Wang, S.; Ghedin, E.; Spiro, D.J.; Saif, L.J. Molecular characterization of a new species in the genus Alphacoronavirus associated with mink epizootic catarrhal gastroenteritis. J. Gen. Virol. 2011, 92, 1369–1379. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Teeling, E.C.; Springer, M.S.; Madsen, O.; Bates, P.; O’Brien, S.J.; Murphy, W.J. A molecular phylogeny for bats illuminates biogeography and the fossil record. Science 2005, 307, 580–584. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.F.; Cowled, C. Bats and Viruses: A New Frontior of Emerging Infectious Diseases; John Wiley Sons Inc.: Hoboken, NJ, USA, 2015. [Google Scholar]
- Yang, X.L.; Tan, C.W.; Anderson, D.E.; Jiang, R.D.; Li, B.; Zhang, W.; Zhu, Y.; Lim, X.F.; Zhou, P.; Liu, X.L.; et al. Characterization of a filovirus (Mengla virus) from Rousettus bats in China. Nat. Microbiol. 2019. [Google Scholar] [CrossRef] [PubMed]
- Olival, K.J.; Hosseini, P.R.; Zambrana-Torrelio, C.; Ross, N.; Bogich, T.L.; Daszak, P. Host and viral traits predict zoonotic spillover from mammals. Nature 2017, 546, 646–650. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Guan, Y.; Zheng, B.J.; He, Y.Q.; Liu, X.L.; Zhuang, Z.X.; Cheung, C.L.; Luo, S.W.; Li, P.H.; Zhang, L.J.; Guan, Y.J.; et al. Isolation and characterization of viruses related to the SARS coronavirus from animals in southern China. Science 2003, 302, 276–278. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.P.; Woo, P.C.Y.; Li, K.S.M.; Huang, Y.; Tsoi, H.W.; Wong, B.H.L.; Wong, S.S.Y.; Leung, S.Y.; Chan, K.H.; Yuen, K.Y. Severe acute respiratory syndrome coronavirus-like virus in Chinese horseshoe bats. Proc. Natl. Acad. Sci. USA 2005, 102, 14040–14045. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Li, W.D.; Shi, Z.L.; Yu, M.; Ren, W.Z.; Smith, C.; Epstein, J.H.; Wang, H.Z.; Crameri, G.; Hu, Z.H.; Zhang, H.J.; et al. Bats are natural reservoirs of SARS-like coronaviruses. Science 2005, 310, 676–679. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.Y.; Li, J.L.; Yang, X.L.; Chmura, A.A.; Zhu, G.J.; Epstein, J.H.; Mazet, J.K.; Hu, B.; Zhang, W.; Peng, C.; et al. Isolation and characterization of a bat SARS-like coronavirus that uses the ACE2 receptor. Nature 2013, 503, 535. [Google Scholar] [CrossRef] [PubMed]
- Lacroix, A.; Duong, V.; Hul, V.; San, S.; Davun, H.; Omaliss, K.; Chea, S.; Hassanin, A.; Theppangna, W.; Silithammavong, S.; et al. Genetic diversity of coronaviruses in bats in Lao PDR and Cambodia. Infect. Genet. Evol. 2017, 48, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Shi, M.; Chommanard, C.; Queen, K.; Zhang, J.; Markotter, W.; Kuzmin, I.V.; Holmes, E.C.; Tong, S. Surveillance of Bat Coronaviruses in Kenya Identifies Relatives of Human Coronaviruses NL63 and 229E and Their Recombination History. J. Virol. 2017, 91, e01953-16. [Google Scholar] [CrossRef] [PubMed]
- Leopardi, S.; Holmes, E.C.; Gastaldelli, M.; Tassoni, L.; Priori, P.; Scaravelli, D.; Zamperin, G.; De Benedictis, P. Interplay between co-divergence and cross-species transmission in the evolutionary history of bat coronaviruses. Infect. Genet. Evol. 2018, 58, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.P.; Li, K.S.M.; Tsang, A.K.L.; Shek, C.T.; Wang, M.; Choi, G.K.Y.; Guo, R.T.; Wong, B.H.L.; Poon, R.W.S.; Lam, C.S.F.; et al. Recent Transmission of a Novel Alphacoronavirus, Bat Coronavirus HKU10, from Leschenault’s Rousettes to Pomona Leaf-Nosed Bats: First Evidence of Interspecies Transmission of Coronavirus between Bats of Different Suborders. J. Virol. 2012, 86, 11906–11918. [Google Scholar] [CrossRef] [PubMed]
- He, B.; Zhang, Y.; Xu, L.; Yang, W.; Yang, F.; Feng, Y.; Xia, L.; Zhou, J.; Zhen, W.; Feng, Y.; et al. Identification of diverse alphacoronaviruses and genomic characterization of a novel severe acute respiratory syndrome-like coronavirus from bats in China. J. Virol. 2014, 88, 7070–7082. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Z.Q.; Chen, D.H.; Tan, W.P.; Qiu, S.Y.; Xu, D.; Liang, H.X.; Chen, M.X.; Li, X.; Lin, Z.S.; Liu, W.K.; et al. Epidemiology and clinical characteristics of human coronaviruses OC43, 229E, NL63, and HKU1: A study of hospitalized children with acute respiratory tract infection in Guangzhou, China. Eur. J. Clin. Microbiol. Infect. Dis. 2018, 37, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Yip, C.C.; Lam, C.S.; Luk, H.K.; Wong, E.Y.; Lee, R.A.; So, L.Y.; Chan, K.H.; Cheng, V.C.; Yuen, K.Y.; Woo, P.C.; et al. A six-year descriptive epidemiological study of human coronavirus infections in hospitalized patients in Hong Kong. Virol. Sin. 2016, 31, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Lin, X.D.; Guo, W.P.; Zhou, R.H.; Wang, M.R.; Wang, C.Q.; Ge, S.; Mei, S.H.; Li, M.H.; Shi, M.; et al. Discovery, diversity and evolution of novel coronaviruses sampled from rodents in China. Virology 2015, 474, 19–27. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Provacia, L.B.; Smits, S.L.; Martina, B.E.; Raj, V.S.; Doel, P.V.; Amerongen, G.V.; Moorman-Roest, H.; Osterhaus, A.D.; Haagmans, B.L. Enteric coronavirus in ferrets, The Netherlands. Emerg. Infect. Dis. 2011, 17, 1570–1571. [Google Scholar] [CrossRef] [PubMed]
- Poon, L.L.; Chu, D.K.; Chan, K.H.; Wong, O.K.; Ellis, T.M.; Leung, Y.H.; Lau, S.K.; Woo, P.C.; Suen, K.Y.; Yuen, K.Y.; et al. Identification of a novel coronavirus in bats. J. Virol. 2005, 79, 2001–2009. [Google Scholar] [CrossRef] [PubMed]
- Chu, D.K.; Poon, L.L.; Chan, K.H.; Chen, H.; Guan, Y.; Yuen, K.Y.; Peiris, J.S. Coronaviruses in bent-winged bats (Miniopterus spp.). J. Gen. Virol. 2006, 87, 2461–2466. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chu, D.K.; Peiris, J.S.; Chen, H.; Guan, Y.; Poon, L.L. Genomic characterizations of bat coronaviruses (1A, 1B and HKU8) and evidence for co-infections in Miniopterus bats. J. Gen. Virol. 2008, 89, 1282–1287. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Du, J.; Yang, L.; Ren, X.; Zhang, J.; Dong, J.; Sun, L.; Zhu, Y.; Yang, F.; Zhang, S.; Wu, Z.; et al. Genetic diversity of coronaviruses in Miniopterus fuliginosus bats. Sci. China Life Sci. 2016, 59, 604–614. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Liang, J.; Yang, X.L.; Li, B.; Liu, Q.; Zhang, Q.; Liu, H.; Kan, H.P.; Wong, K.C.; Chek, S.N.; He, X.; et al. Detection of diverse viruses in alimentary specimens of bats in Macau. Virol. Sin. 2017, 32, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.D.; Wang, W.; Hao, Z.Y.; Wang, Z.X.; Guo, W.P.; Guan, X.Q.; Wang, M.R.; Wang, H.W.; Zhou, R.H.; Li, M.H.; et al. Extensive diversity of coronaviruses in bats from China. Virology 2017, 507, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Woo, P.C.; Lau, S.K.; Li, K.S.; Poon, R.W.; Wong, B.H.; Tsoi, H.W.; Yip, B.C.; Huang, Y.; Chan, K.H.; Yuen, K.Y. Molecular diversity of coronaviruses in bats. Virology 2006, 351, 180–187. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Woo, P.C.; Wang, M.; Lau, S.K.; Xu, H.; Poon, R.W.; Guo, R.; Wong, B.H.; Gao, K.; Tsoi, H.W.; Huang, Y.; et al. Comparative analysis of twelve genomes of three novel group 2c and group 2d coronaviruses reveals unique group and subgroup features. J. Virol. 2007, 81, 1574–1585. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.; Feng, Y.; Chen, H.; Luk, H.K.; Yang, W.H.; Li, K.S.; Zhang, Y.Z.; Huang, Y.; Song, Z.Z.; Chow, W.N.; et al. Severe Acute Respiratory Syndrome (SARS) Coronavirus ORF8 Protein Is Acquired from SARS-Related Coronavirus from Greater Horseshoe Bats through Recombination. J. Virol. 2015, 89, 10532–10547. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Xu, L.; Zhang, F.; Yang, W.; Jiang, T.; Lu, G.; He, B.; Li, X.; Hu, T.; Chen, G.; Feng, Y.; et al. Detection and characterization of diverse alpha- and betacoronaviruses from bats in China. Virol. Sin. 2016, 31, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Hu, R.; Tang, X.; Wu, C.; He, Q.; Zhao, Z.; Chen, H.; Wu, B. Occurrence and investigation of enteric viral infections in pigs with diarrhea in China. Arch. Virol. 2013, 158, 1631–1636. [Google Scholar] [CrossRef] [PubMed]
- Tang, X.C.; Zhang, J.X.; Zhang, S.Y.; Wang, P.; Fan, X.H.; Li, L.F.; Li, G.; Dong, B.Q.; Liu, W.; Cheung, C.L.; et al. Prevalence and genetic diversity of coronaviruses in bats from China. J. Virol. 2006, 80, 7481–7490. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.; Woo, P.C.; Li, K.S.; Huang, Y.; Wang, M.; Lam, C.S.; Xu, H.; Guo, R.; Chan, K.H.; Zheng, B.J.; et al. Complete genome sequence of bat coronavirus HKU2 from Chinese horseshoe bats revealed a much smaller spike gene with a different evolutionary lineage from the rest of the genome. Virology 2007, 367, 428–439. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lau, S.K.; Li, K.S.; Huang, Y.; Shek, C.T.; Tse, H.; Wang, M.; Choi, G.K.; Xu, H.; Lam, C.S.; Guo, R.; et al. Ecoepidemiology and complete genome comparison of different strains of severe acute respiratory syndrome-related Rhinolophus bat coronavirus in China reveal bats as a reservoir for acute, self-limiting infection that allows recombination events. J. Virol. 2010, 84, 2808–2819. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.; Woo, P.C.; Li, K.S.; Tsang, A.K.; Fan, R.Y.; Luk, H.K.; Cai, J.P.; Chan, K.H.; Zheng, B.J.; Wang, M.; et al. Discovery of a novel coronavirus, China Rattus coronavirus HKU24, from Norway rats supports the murine origin of Betacoronavirus 1 and has implications for the ancestor of Betacoronavirus lineage A. J. Virol. 2015, 89, 3076–3092. [Google Scholar] [CrossRef] [PubMed]
- Bardos, V.; Schwanzer, V.; Pesko, J. Identification of Tettnang virus (‘possible arbovirus’) as mouse hepatitis virus. Intervirology 1980, 13, 275–283. [Google Scholar] [PubMed]
- Corman, V.M.; Kallies, R.; Philipps, H.; Gopner, G.; Muller, M.A.; Eckerle, I.; Brunink, S.; Drosten, C.; Drexler, J.F. Characterization of a novel betacoronavirus related to middle East respiratory syndrome coronavirus in European hedgehogs. J. Virol. 2014, 88, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.M.; Wang, N.; Yang, X.L.; Liu, H.Z.; Zhang, W.; Li, B.; Hu, B.; Peng, C.; Geng, Q.B.; Zhu, G.J.; et al. Discovery of Novel Bat Coronaviruses in South China That Use the Same Receptor as Middle East Respiratory Syndrome Coronavirus. J. Virol. 2018, 92, e00116-18. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.P.; Zhang, L.B.; Luk, H.K.H.; Xiong, L.F.; Peng, X.W.; Li, K.S.M.; He, X.Y.; Zhao, P.S.H.; Fan, R.Y.Y.; Wong, A.C.P.; et al. Receptor Usage of a Novel Bat Lineage C Betacoronavirus Reveals Evolution of Middle East Respiratory Syndrome-Related Coronavirus Spike Proteins for Human Dipeptidyl Peptidase 4 Binding. J. Infect. Dis. 2018, 218, 197–207. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.P.; Li, K.S.M.; Tsang, A.K.L.; Lam, C.S.F.; Ahmed, S.; Chen, H.L.; Chan, K.H.; Woo, P.C.Y.; Yuen, K.Y. Genetic Characterization of Betacoronavirus Lineage C Viruses in Bats Reveals Marked Sequence Divergence in the Spike Protein of Pipistrellus Bat Coronavirus HKU5 in Japanese Pipistrelle: Implications for the Origin of the Novel Middle East Respiratory Syndrome Coronavirus. J. Virol. 2013, 87, 8638–8650. [Google Scholar] [PubMed][Green Version]
- Yang, L.; Wu, Z.Q.; Ren, X.W.; Yang, F.; Zhang, J.P.; He, G.M.; Dong, J.; Sun, L.L.; Zhu, Y.F.; Zhang, S.Y.; et al. MERS-Related Betacoronavirus in Vespertilio superans Bats, China. Emerg. Infect. Dis. 2014, 20, 1260–1262. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Liu, W.J.; Xu, W.; Jin, T.; Zhao, Y.; Song, J.; Shi, Y.; Ji, W.; Jia, H.; Zhou, Y.; et al. A Bat-Derived Putative Cross-Family Recombinant Coronavirus with a Reovirus Gene. PLoS Pathog. 2016, 12, e1005883. [Google Scholar] [CrossRef] [PubMed]
- Obameso, J.O.; Li, H.; Jia, H.; Han, M.; Zhu, S.; Huang, C.; Zhao, Y.; Zhao, M.; Bai, Y.; Yuan, F.; et al. The persistent prevalence and evolution of cross-family recombinant coronavirus GCCDC1 among a bat population: A two-year follow-up. Sci. China Life Sci. 2017, 60, 1357–1363. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Li, B.; Jiang, R.D.; Hu, B.J.; Luo, D.S.; Zhu, G.J.; Hu, B.; Liu, H.Z.; Zhang, Y.Z.; Yang, X.L.; et al. Longitudinal Surveillance of Betacoronaviruses in Fruit Bats in Yunnan Province, China During 2009–2016. Virol. Sin. 2018, 33, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Lau, S.K.; Poon, R.W.; Wong, B.H.; Wang, M.; Huang, Y.; Xu, H.; Guo, R.; Li, K.S.; Gao, K.; Chan, K.H.; et al. Coexistence of different genotypes in the same bat and serological characterization of Rousettus bat coronavirus HKU9 belonging to a novel Betacoronavirus subgroup. J. Virol. 2010, 84, 11385–11394. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.; Li, Y.; Yang, X.; Zhang, H.; Zhou, P.; Zhang, Y.; Shi, Z. Metagenomic analysis of viruses from bat fecal samples reveals many novel viruses in insectivorous bats in China. J. Virol. 2012, 86, 4620–4630. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Li, W.D.; Yu, M.; Hao, P.; Zhang, Y.; Zhou, P.; Zhang, S.Y.; Zhao, G.P.; Zhong, Y.; Wang, S.Y.; et al. Full-length genome sequences of two SARS-like coronaviruses in horseshoe bats and genetic variation analysis. J. Gen. Virol. 2006, 87, 3355–3359. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yuan, J.F.; Hon, C.C.; Li, Y.; Wang, D.M.; Xu, G.L.; Zhang, H.J.; Zhou, P.; Poon, L.L.M.; Lam, T.T.Y.; Leung, F.C.C.; et al. Intraspecies diversity of SARS-like coronaviruses in Rhinolophus sinicus and its implications for the origin of SARS coronaviruses in humans. J. Gen. Virol. 2010, 91, 1058–1062. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.G.; Yang, L.; Ren, X.W.; Zhang, J.P.; Yang, F.; Zhang, S.Y.; Jin, Q. ORF8-Related Genetic Evidence for Chinese Horseshoe Bats as the Source of Human Severe Acute Respiratory Syndrome Coronavirus. J. Infect. Dis. 2016, 213, 579–583. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.L.; Hu, B.; Wang, B.; Wang, M.N.; Zhang, Q.; Zhang, W.; Wu, L.J.; Ge, X.Y.; Zhang, Y.Z.; Daszak, P.; et al. Isolation and Characterization of a Novel Bat Coronavirus Closely Related to the Direct Progenitor of Severe Acute Respiratory Syndrome Coronavirus. J. Virol. 2016, 90, 3253–3256. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Zeng, L.P.; Yang, X.L.; Ge, X.Y.; Zhang, W.; Li, B.; Xie, J.Z.; Shen, X.R.; Zhang, Y.Z.; Wang, N.; et al. Discovery of a rich gene pool of bat SARS-related coronaviruses provides new insights into the origin of SARS coronavirus. PLoS Pathog. 2017, 13, e1006698. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Zhu, C.Q.; Ai, L.L.; He, T.; Wang, Y.; Ye, F.Q.; Yang, L.; Ding, C.X.; Zhu, X.H.; Lv, R.C.; et al. Genomic characterization and infectivity of a novel SARS-like coronavirus in Chinese bats. Emerg. Microbes Infect. 2018, 7, 154. [Google Scholar] [CrossRef] [PubMed]
- Hu, D.; Zhu, C.; Wang, Y.; Ai, L.; Yang, L.; Ye, F.; Ding, C.; Chen, J.; He, B.; Zhu, J.; et al. Virome analysis for identification of novel mammalian viruses in bats from Southeast China. Sci. Rep. 2017, 7, 10917. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Woo, P.C.; Lau, S.K.; Lam, C.S.; Lau, C.C.; Tsang, A.K.; Lau, J.H.; Bai, R.; Teng, J.L.; Tsang, C.C.; Wang, M.; et al. Discovery of seven novel Mammalian and avian coronaviruses in the genus deltacoronavirus supports bat coronaviruses as the gene source of alphacoronavirus and betacoronavirus and avian coronaviruses as the gene source of gammacoronavirus and deltacoronavirus. J. Virol. 2012, 86, 3995–4008. [Google Scholar] [PubMed]
- Woo, P.C.; Lau, S.K.; Tsang, C.C.; Lau, C.C.; Wong, P.C.; Chow, F.W.; Fong, J.Y.; Yuen, K.Y. Coronavirus HKU15 in respiratory tract of pigs and first discovery of coronavirus quasispecies in 5′-untranslated region. Emerg. Microbes Infect. 2017, 6, e53. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Qu, X.; Li, W.; Han, Z.; Yu, M.; Zhou, P.; Zhang, S.Y.; Wang, L.F.; Deng, H.; Shi, Z. Difference in receptor usage between severe acute respiratory syndrome (SARS) coronavirus and SARS-like coronavirus of bat origin. J. Virol. 2008, 82, 1899–1907. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Wu, Z.Q.; Ren, X.W.; Yang, F.; He, G.M.; Zhang, J.P.; Dong, J.; Sun, L.L.; Zhu, Y.F.; Du, J.; et al. Novel SARS-like Betacoronaviruses in Bats, China, 2011. Emerg. Infect. Dis. 2013, 19, 989–991. [Google Scholar] [CrossRef] [PubMed]
- Menachery, V.D.; Yount, B.L., Jr.; Debbink, K.; Agnihothram, S.; Gralinski, L.E.; Plante, J.A.; Graham, R.L.; Scobey, T.; Ge, X.Y.; Donaldson, E.F.; et al. A SARS-like cluster of circulating bat coronaviruses shows potential for human emergence. Nat. Med. 2015, 21, 1508–1513. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, N.; Li, S.Y.; Yang, X.L.; Huang, H.M.; Zhang, Y.J.; Guo, H.; Luo, C.M.; Miller, M.; Zhu, G.; Chmura, A.A.; et al. Serological Evidence of Bat SARS-Related Coronavirus Infection in Humans, China. Virol. Sin. 2018, 33, 104–107. [Google Scholar] [CrossRef] [PubMed]
- Reusken, C.B.; Raj, V.S.; Koopmans, M.P.; Haagmans, B.L. Cross host transmission in the emergence of MERS coronavirus. Curr. Opin. Virol. 2016, 16, 55–62. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Corman, V.M.; Ithete, N.L.; Richards, L.R.; Schoeman, M.C.; Preiser, W.; Drosten, C.; Drexler, J.F. Rooting the phylogenetic tree of middle East respiratory syndrome coronavirus by characterization of a conspecific virus from an African bat. J. Virol. 2014, 88, 11297–11303. [Google Scholar] [CrossRef] [PubMed]
- Anthony, S.J.; Gilardi, K.; Menachery, V.D.; Goldstein, T.; Ssebide, B.; Mbabazi, R.; Navarrete-Macias, I.; Liang, E.; Wells, H.; Hicks, A.; et al. Further Evidence for Bats as the Evolutionary Source of Middle East Respiratory Syndrome Coronavirus. MBio 2017, 8, e00373-17. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Du, L.; Liu, C.; Wang, L.; Ma, C.; Tang, J.; Baric, R.S.; Jiang, S.; Li, F. Receptor usage and cell entry of bat coronavirus HKU4 provide insight into bat-to-human transmission of MERS coronavirus. Proc. Natl. Acad. Sci. USA 2014, 111, 12516–12521. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wang, L.; Su, S.; Bi, Y.; Wong, G.; Gao, G.F. Bat-Origin Coronaviruses Expand Their Host Range to Pigs. Trends Microbiol. 2018, 26, 466–470. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Tachedjian, M.; Wynne, J.W.; Boyd, V.; Cui, J.; Smith, I.; Cowled, C.; Ng, J.H.; Mok, L.; Michalski, W.P.; et al. Contraction of the type I IFN locus and unusual constitutive expression of IFN-alpha in bats. Proc. Natl. Acad. Sci. USA 2016, 113, 2696–2701. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Li, Y.; Shen, X.; Goh, G.; Zhu, Y.; Cui, J.; Wang, L.F.; Shi, Z.L.; Zhou, P. Dampened STING-Dependent Interferon Activation in Bats. Cell Host Microbe 2018, 23, 297–301. [Google Scholar] [CrossRef] [PubMed]
Figure 1.Geographical distribution of bat coronaviruses (CoVs) and their corresponding bat hosts in China. Each red box represents one CoV positive sample found in that particular bat species. One dot matrix was drawn for each province where a CoV positive sample had been reported. Guangdong Province, where SARS and SADS began, is circled in red. Abbreviations of bat species and virus species are indicated.
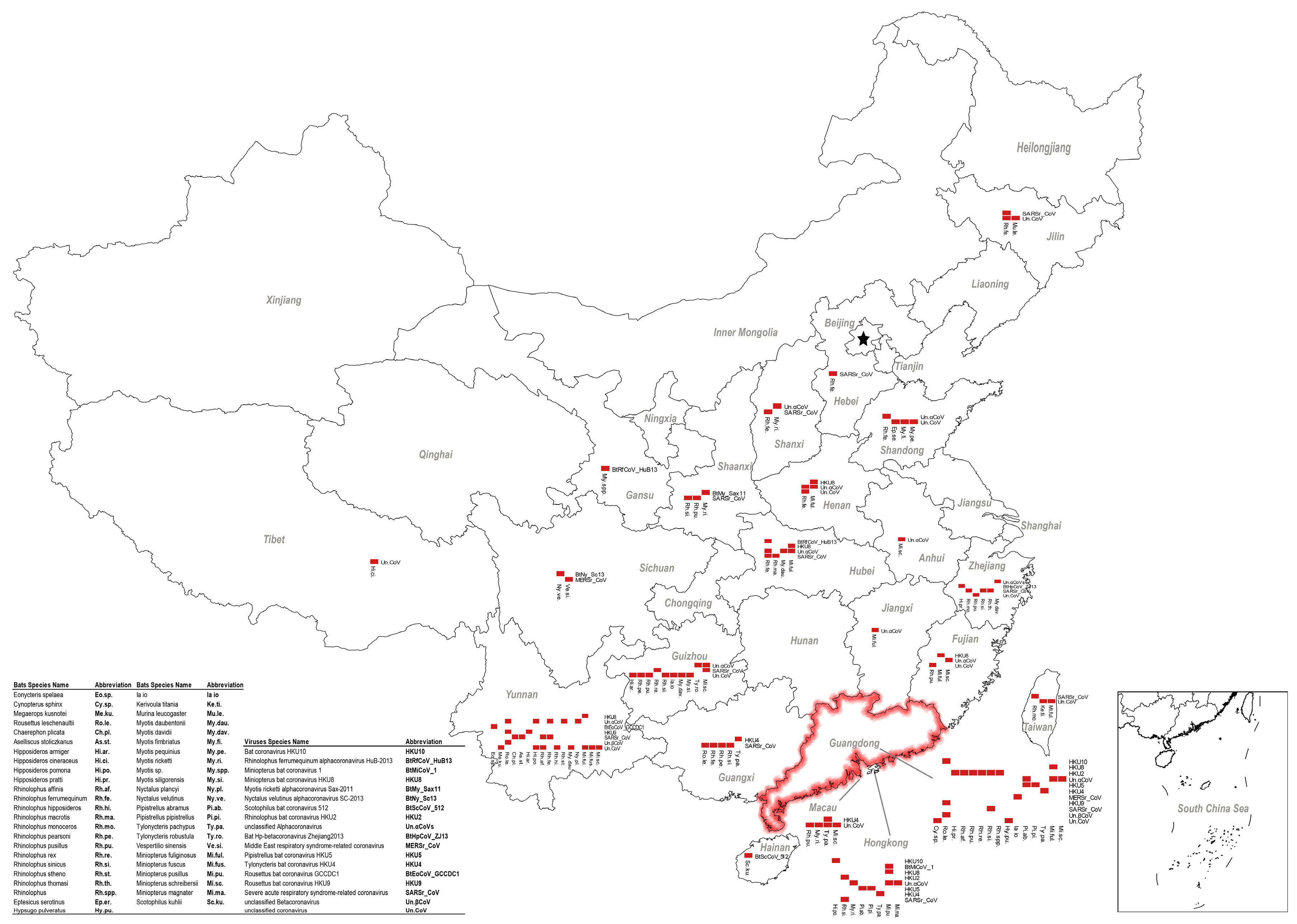
Figure 2.Genetic diversity of bat CoV in China. Sequences of 440 bp conserved the viral polymerase (RdRp) region for each CoV species were compared to related reference sequences. Reference genomes used: BtCoV-HKU10, NC_018871.1; BtRfCoV-HuB13, NC_028814.1; BtMiCoV-1, EU420138.1; BtMiCoV-HKU8, NC_010438.1; BtRhCoV-HKU2, MF094682.1; BtHpCoV-ZJ13, NC_025217.1; MERSr-CoV, NC_038294.1; BtPiCoV-HKU5, NC_009020.1; BtTyCoV-HKU4, NC_009019.1; BtRoCoV-GCCDC1, MG762606.1; BtRoCoV-HKU9, NC_009021.1; and SARSr-CoV, NC_004718.3. Notably, samples that were positive for BtMy-Sax11, BtNy-Sc13, and BtScCoV-512 were also identified in China. These were not taken into account here as too few sequences were available.
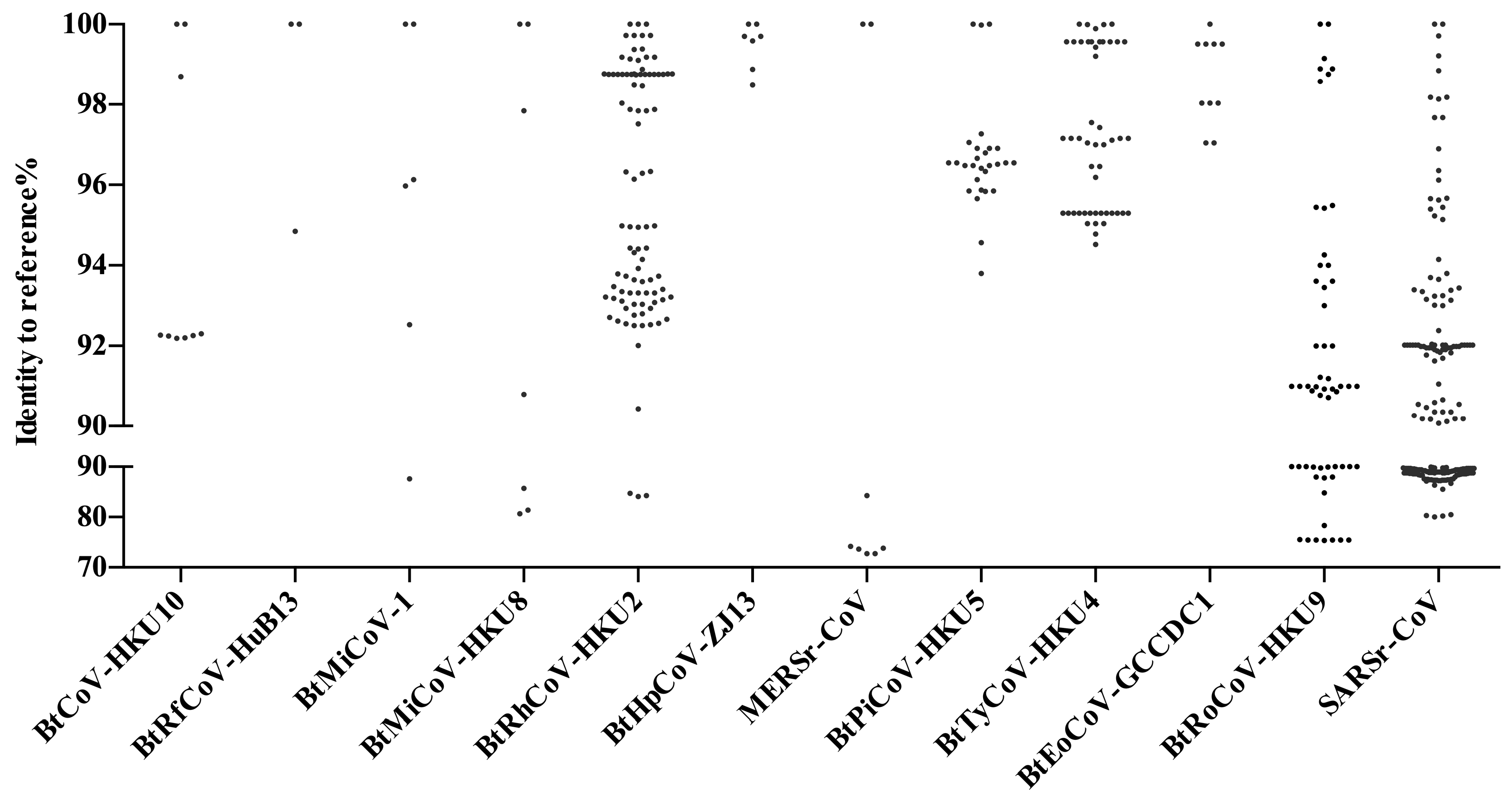
Table 1.International Committee of Taxonomy of Viruses (ICTV) classification of coronaviruses species, reservoir hosts, and presence reported in China.
| Coronavirus Species | Abbreviations | Human | Bats | Other Animals | Reported in China | |
|---|---|---|---|---|---|---|
| Bat coronavirus HKU10 | BtCoV-HKU10 | Yes | Yes [7,8,26,27] | α-CoV | ||
| Bat coronavirus CDPHE15 | BtCoV-CDPHE15 | Yes | No | |||
| Rhinolophus ferrumequinum alphacoronavirus HuB-2013 | BtRfCoV-HuB13 | Yes | Yes [8] | |||
| * Human coronavirus 229E | HCoV-229E | Yes | Yes [28,29] | |||
| Lucheng Rn rat coronavirus | LRNV | Yes (rat) | Yes [30] | |||
| Ferret coronavirus | FRCoV | Yes (ferret) | No [31] | |||
| * Mink coronavirus 1 | MCoV | Yes (mink) | No [14] | |||
| Miniopterus bat coronavirus 1 | BtMiCoV-1 | Yes | Yes [7,8,32,33,34,35,36,37] | |||
| Miniopterus bat coronavirus HKU8 | BtMiCoV-HKU8 | Yes | Yes [7,8,33,34,35,37,38,39,40,41] | |||
| Myotis ricketti alphacoronavirus Sax-2011 | BtMy-Sax11 | Yes | Yes [8,37] | |||
| Nyctalus velutinus alphacoronavirus SC-2013 | BtNy-Sc13 | Yes | Yes [8] | |||
| * Porcine epidemic diarrhea virus | PEDV | Yes (pig) | Yes [42] | |||
| Scotophilus bat coronavirus 512 | BtScCoV-512 | Yes | Yes [37] | |||
| * Rhinolophus bat coronavirus HKU2 (SADS) | BtRhCoV-HKU2 | Yes | Yes | Yes [2,7,8,38,43,44,45] | ||
| * Human coronavirus NL63 | HCoV-NL63 | Yes | Yes [28,29] | |||
| NL63-related bat coronavirus strain BtKYNL63-9b | BtKYNL63 | Yes | No [24] | |||
| * Alphacoronavirus 1 (Transmissible gastroenteritis virus) | TGEV | Yes (pig) | Yes [42] | |||
| China Rattus coronavirus HKU24 | RtCoV-HKU24 | Yes (rat) | Yes [46] | β-CoV | ||
| * Human coronavirus HKU1 | HCoV-HKU1 | Yes | Yes [28,29] | |||
| * Murine coronavirus (Murine hepatitis coronavirus) | MHV | Yes (mouse) | No [47] | |||
| Bat Hp-betacoronavirus Zhejiang2013 | BtHpCoV-ZJ13 | Yes | Yes [8] | |||
| Hedgehog coronavirus 1 | EriCoV-1 | Yes (hedgehog) | No [48] | |||
| * Middle East respiratory syndrome-related coronavirus | MERSr-CoV | Yes | Yes | Yes [49,50] | ||
| Pipistrellus bat coronavirus HKU5 | BtPiCoV-HKU5 | Yes | Yes [38,39,49,51,52] | |||
| Tylonycteris bat coronavirus HKU4 | BtTyCoV-HKU4 | Yes | Yes [36,38,39,49,50,51] | |||
| Rousettus bat coronavirus GCCDC1 | # BtEoCoV-GCCDC1 | Yes | Yes [53,54,55] | |||
| Rousettus bat coronavirus HKU9 | BtRoCoV-HKU9 | Yes | Yes [39,55,56,57] | |||
| * Severe acute respiratory syndrome-related coronavirus | SARSr-CoV | Yes | Yes | Yes [7,8,20,21,22,27,37,40,45,58,59,60,61,62,63,64] | ||
| * Betacoronavirus 1 (Human coronavirus OC43) | HCoV-OC43 | Yes | Yes [28,29] | |||
| Wigeon coronavirus HKU20 | WiCoV-HKU20 | Yes (bird) | Yes [65] | δ-CoV | ||
| Bulbul coronavirus HKU11 | BuCoV-HKU11 | Yes (bird) | Yes [65] | |||
| Coronavirus HKU15 | PoCoV-HKU15 | Yes (pig) | Yes [66] | |||
| Munia coronavirus HKU13 | MuCoV-HKU13 | Yes (bird) | Yes [65] | |||
| White-eye coronavirus HKU16 | WECoV-HKU13 | Yes (bird) | Yes [65] | |||
| Night heron coronavirus HKU19 | NHCoV-HKU19 | Yes (bird) | Yes [65] | |||
| Common moorhen coronavirus HKU21 | CMCoV-HKU21 | Yes (bird) | Yes [65] | |||
| *? Beluga whale coronavirus SW1 | BWCoV-SW1 | Yes (whale) | No [13] | γ-CoV | ||
| * Avian infectious bronchitis virus | IBV | Yes (bird) | Yes [12] |
* The disease-causing CoVs are indicated and the three zoonotic CoVs are in bold. *? BWCoV-SW1 was found in a sick whale, but whether it was the etiological agent of the infection was not proven. # Carrier of this virus was confirmed as Eonycteris spelaea, but not Rousettus bats. The virus was renamed accordingly.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Johns Hopkins Expert: Use Antibodies As Temporary Solution To Coronavirus


BALTIMORE (WJZ) — Johns Hopkins is also on the frontlines of the coronavirus pandemic.
 © Provided by CBS Baltimore
© Provided by CBS Baltimore
Infectious disease experts at Johns Hopkins have a possible solution to keeping COVID-19 under control.
They said the U.S. should considering using the antibodies of people who have recovered from the infection to treat patients and protect healthcare workers.
CORONAVIRUS COVERAGE:
- What Is A Coronavirus?
- The Symptoms Of Coronavirus And What You Should Do If You Feel Sick
- LIST: EPA Releases Names Of Disinfectants You Can Use Against Coronavirus
- Coronavirus-Related Scams Are Going Around. Here’s What To Watch Out For
- What We Know About Coronavirus In Maryland
- Coronavirus Closings: Baltimore, Maryland Events Postponed Or Canceled
- Latest coronavirus stories from WJZ
They said using an antibody that contains serum from recovered patients has a long history of being a temporary solution against infectious diseases.
Already, more than half of those infected by coronavirus worldwide have recovered.
For the latest information on coronavirus go to the Maryland Health Department’s website or call 211. You can find all of WJZ’s coverage oncoronavirus in Maryland here.
No comments:
Post a Comment